
21 Plant Transport
At the end of this chapter you will be able to:
- Identify the different tissue types in plants
- Explain how water moves from the roots of a plant up to the leaves
- Explain how food made by the plant moves from its leaves to the roots for storage
- Define water potential and explain how it is influenced by solutes, pressure, gravity, and the matric potential
- Describe how water potential, evapotranspiration, and stomatal regulation influence how water is transported in plants
- Compare and contrast xylem and phloem
- Define water potential and explain how it is influenced by solutes and pressure
Introduction
All living organisms need a way to move nutrients, gasses, water, and waste within their bodies. Some plants and animals are small enough and/or thin enough that they can rely on diffusion for this process; however, larger plants and animals need a more organized system to transport things from one part of their body to another. A common analogy is to consider cells acting as “pipes” for such transport. Before we begin to discuss transport in more detail, we will first need to gain some understanding of plant tissues in general.

Plant Tissues
Plants are multicellular eukaryotes with tissue systems made of various cell types that carry out specific functions. Plant tissue systems fall into one of two general types: meristematic tissue and permanent (or non-meristematic) tissue. Cells of the meristematic tissue are found in meristems, which are plant regions of continuous cell division and growth. Meristematic tissue cells are either undifferentiated or incompletely differentiated, and they continue to divide and contribute to the growth of the plant. In contrast, permanent tissue consists of plant cells that are no longer actively dividing.
Meristematic tissues consist of three types, based on their location in the plant. Apical meristems contain meristematic tissue located at the tips of stems and roots, which enable a plant to extend in length. Lateral meristems facilitate growth in thickness or girth in a maturing plant. Intercalary meristems occur only in monocots, at the bases of leaf blades and at nodes (the areas where leaves attach to a stem). This tissue enables the monocot leaf blade to increase in length from the leaf base; for example, it allows lawn grass leaves to elongate even after repeated mowing.
Meristems produce cells that quickly differentiate, or specialize, and become permanent tissue. Such cells take on specific roles and lose their ability to divide further. They differentiate into three main types: dermal, vascular, and ground tissue. Dermal tissue covers and protects the plant, and vascular tissue transports water, minerals, and sugars to different parts of the plant. Ground tissue serves as a site for photosynthesis, provides a supporting matrix for the vascular tissue, and helps to store water and sugars.
Secondary tissues are either simple (composed of similar cell types) or complex (composed of different cell types). Dermal tissue, for example, is a simple tissue that covers the outer surface of the plant and controls gas exchange. Vascular tissue is an example of a complex tissue, and is made of two specialized conducting tissues: xylem and phloem. Xylem tissue transports water and nutrients from the roots to different parts of the plant, and includes three different cell types: vessel elements and tracheids (both of which conduct water), and xylem parenchyma. Phloem tissue, which transports organic compounds from the site of photosynthesis to other parts of the plant, consists of four different cell types: sieve cells (which conduct photosynthates), companion cells, phloem parenchyma, and phloem fibers. Unlike xylem conducting cells, phloem conducting cells are alive at maturity. The xylem and phloem always lie adjacent to each other (Fig 1). In stems, the xylem and the phloem form a structure called a vascular bundle; in roots, this is termed the vascular stele or vascular cylinder.
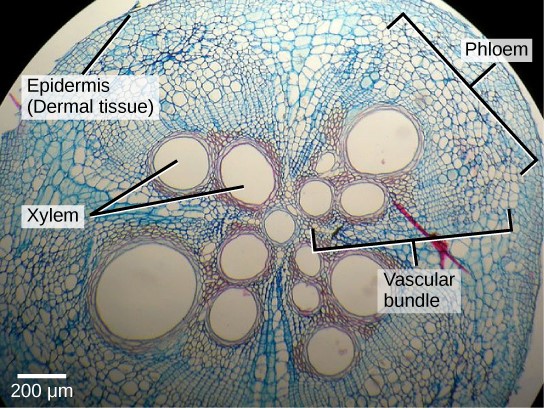
Figure 1: This light micrograph shows a cross section of a squash (Curcurbita maxima) stem. Each teardrop-shaped vascular bundle consists of large xylem vessels toward the inside and smaller phloem cells toward the outside. Xylem cells, which transport water and nutrients from the roots to the rest of the plant, are dead at functional maturity. Phloem cells, which transport sugars and other organic compounds from photosynthetic tissue to the rest of the plant, are living. The vascular bundles are encased in ground tissue and surrounded by dermal tissue. (credit: modification of work by “(biophotos)”/Flickr; scale-bar data from Matt Russell)
Because it is important to understand plant transport, we will focus more on the vascular tissue of plants: xylem and phloem. These tissues make up the vascular tissue of the stem and are arranged in distinct strands called vascular bundles, which run up and down the length of the stem. When the stem is viewed in cross-section, the vascular bundles of dicot stems are arranged in a ring. In plants with stems that live for more than one year, the individual bundles grow together and produce the characteristic growth rings. In monocot stems, the vascular bundles are randomly scattered throughout the ground tissue (Fig 2).

Figure 2: In (a) dicot stems, vascular bundles are arranged around the periphery of the ground tissue. The xylem tissue is located toward the interior of the vascular bundle, and phloem is located toward the exterior. Sclerenchyma fibers cap the vascular bundles. In (b) monocot stems, vascular bundles composed of xylem and phloem tissues are scattered throughout the ground tissue.
Xylem tissue has three types of cells: xylem parenchyma, vessel elements (Figs 3 & 4) and tracheids (Fig 5). The latter two types conduct water and are dead at maturity. Tracheids are xylem cells with thick secondary cell walls that are lignified. Water moves from one tracheid to another through regions on the side walls known as pits, where secondary walls are absent. Vessel elements are xylem cells with thinner walls; they are shorter than tracheids. Each vessel element is connected to the next by means of a perforation plate at the end walls of the element. Water moves through the perforation plates to travel up the plant.
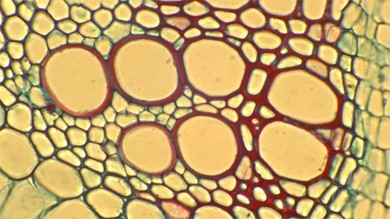
Figure 3: A cross-section micrograph of large xylem vessel element cells with thick red-stained lignified cell walls. Xylem parenchyma adjacent to the cells have blue green cell walls.
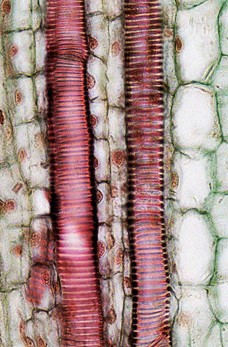

Figure 4: Longitudinal section of xylem vessel element cells with annular lignification in two different species of plants: Coleus (left) and Corn (right).
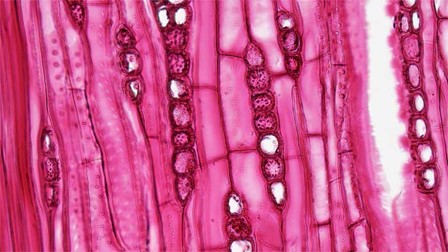
Figure 5: Tracheids in tangential section in Quercus (oak). Long cells with oblique end walls and round pits in the walls are tracheids. Small round cells are lines of ray parenchyma with cellulose cell walls
Phloem tissue is composed of sieve-tube cells, companion cells (Fig 6), phloem parenchyma, and phloem fibers. A series of sieve-tube cells (also called sieve-tube elements) are arranged end to end to make up a long sieve tube, which transports organic substances such as sugars and amino acids. The sugars flow from one sieve-tube cell to the next through perforated sieve plates, which are found at the end junctions between two cells. Although still alive at maturity, the nucleus and other cell components of the sieve-tube cells have disintegrated. Companion cells are found alongside the sieve-tube cells, providing them with metabolic support. The companion cells contain more ribosomes and mitochondria than the sieve-tube cells, which lack some cellular organelles.
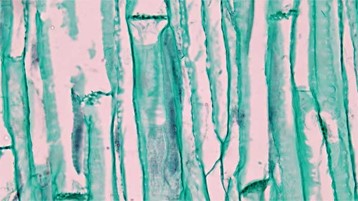
Figure 6: Phloem tissue in longitudinal section including sieve cells and companion cells.
Roots
Another organ of the plant that is important to understand for its part in transport is the root system. The tip of the root is protected by the root cap, a structure exclusive to roots and unlike any other plant structure. The root cap is continuously replaced because it gets damaged easily as the root pushes through soil. The root tip can be divided into three zones: a zone of cell division, a zone of elongation, and a zone of maturation and differentiation. The zone of cell division is closest to the root tip; it is made up of the actively dividing cells of the root meristem. The zone of elongation is where the newly formed cells increase in length, thereby lengthening the root. Beginning at the first root hair is the zone of cell maturation where the root cells begin to differentiate into special cell types. All three zones are in the first centimeter or so of the root tip.
The root has an outer layer of cells called the epidermis, which surrounds areas of ground tissue and vascular tissue. The epidermis provides protection and helps in absorption. Root hairs, which are extensions of root epidermal cells, increase the surface area of the root, greatly contributing to the absorption of water and minerals.
Inside the root, the ground tissue forms two regions: the cortex and the pith (Fig 7). Compared to stems, roots have lots of cortex and little pith. Both regions include cells that store photosynthetic products. The cortex is between the epidermis and the vascular tissue, whereas the pith lies between the vascular tissue and the center of the root.
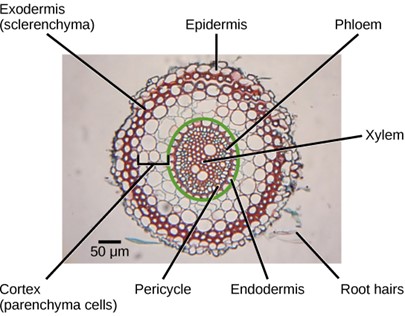
Figure 7: Staining reveals different cell types in this light micrograph of a wheat (Triticum) root cross section. Sclerenchyma cells of the exodermis and xylem cells stain red, and phloem cells stain blue. Other cell types stain black. The stele, or vascular tissue, is the area inside endodermis (indicated by a green ring). Root hairs are visible outside the epidermis. (credit: scale-bar data from Matt Russell)
The vascular tissue in the root is arranged in the inner portion of the root, which is called the stele. A layer of cells known as the endodermis separates the stele from the ground tissue in the outer portion of the root. The endodermis is exclusive to roots and serves as a checkpoint for materials entering the root’s vascular system. A waxy substance called suberin is present on the walls of the endodermal cells. This waxy region, known as the Casparian strip, forces water and solutes to cross the plasma membranes of endodermal cells instead of slipping between the cells. This ensures that only materials required by the root pass through the endodermis, while toxic substances and pathogens are generally excluded. The outermost cell layer of the root’s vascular tissue is the pericycle, an area that can give rise to lateral roots. In dicot roots, the xylem and phloem of the stele are arranged alternately in an X shape, whereas in monocot roots, the vascular tissue is arranged in a ring around the pith (Fig 8).

Figure 8: In (left) typical dicots, the vascular tissue forms an X shape in the center of the root. In (right) typical monocots, the phloem cells and the larger xylem cells form a characteristic ring around the central pith.
Transport of Water in Plants
Transpiration is the loss of water from the plant through evaporation at the leaf surface. It is the main driver of water movement in the xylem. Transpiration is caused by the evaporation of water at the leaf–atmosphere interface; it creates negative pressure (tension) at the leaf surface. Water from the roots is pulled up by this tension. At night, when stomata, the pores in the leaves of plants, shut and transpiration stops, the water is held in the stem and leaf by the adhesion of water to the cell walls of the xylem vessels and tracheids, and the cohesion of water molecules to each other. This is called the cohesion–tension theory of sap ascent.
Inside the leaf at the cellular level, water on the surface of mesophyll cells (general body cells) saturates the cellulose microfibrils of the primary cell wall. The leaf contains many large intercellular air spaces for the exchange of oxygen for carbon dioxide, which is required for photosynthesis. The wet cell wall is exposed to this leaf internal air space, and the water on the surface of the cells evaporates into the air spaces, decreasing the thin film on the surface of the mesophyll cells.
This decrease creates a greater tension on the water in the mesophyll cells (Fig 9), thereby increasing the pull on the water in the xylem vessels. The xylem vessels and tracheids are structurally adapted to cope with large changes in pressure. Rings in the vessels maintain their tubular shape, much like the rings on a vacuum cleaner hose keep the hose open while it is under pressure. Small perforations between vessel elements reduce the number and size of gas bubbles that can form via a process called cavitation. The formation of gas bubbles in xylem interrupts the continuous stream of water from the base to the top of the plant, causing a break termed an embolism in the flow of xylem sap. The taller the tree, the greater the tension forces needed to pull water, and the more cavitation events. In larger trees, the resulting embolisms can plug xylem vessels, making them non-functional. Transpiration is a passive process, meaning that metabolic energy in the form of ATP is not required for water movement. The energy driving transpiration is the difference in energy between the water in the soil and the water in the atmosphere. However, transpiration is tightly controlled.
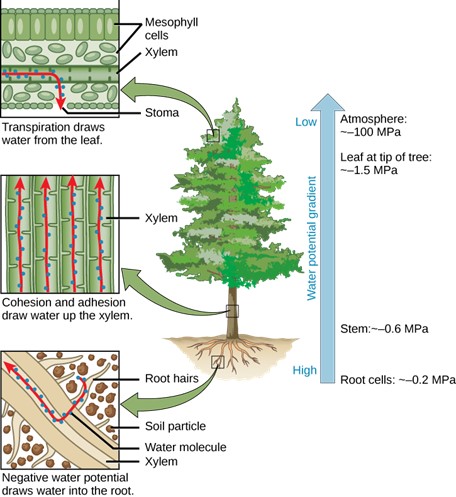
Figure 9: The cohesion–tension theory of sap ascent is shown. Evaporation from the mesophyll cells produces a negative water potential gradient that causes water to move upwards from the roots through the xylem.
The atmosphere to which the leaf is exposed drives transpiration, but also causes massive water loss from the plant. Up to 90 percent of the water taken up by roots may be lost through transpiration.
Leaves are covered by a waxy cuticle on the outer surface that prevents the loss of water. Regulation of transpiration, therefore, is achieved primarily through the opening and closing of stomata on the leaf surface. Stomata are surrounded by two specialized cells called guard cells, which open and close in response to environmental cues such as light intensity and quality, leaf water status, and carbon dioxide concentrations (Fig 10). Stomata must open to allow air containing carbon dioxide and oxygen to diffuse into the leaf for photosynthesis and respiration. When stomata are open, however, water vapor is lost to the external environment, increasing the rate of transpiration. Therefore, plants must maintain a balance between efficient photosynthesis and water loss.
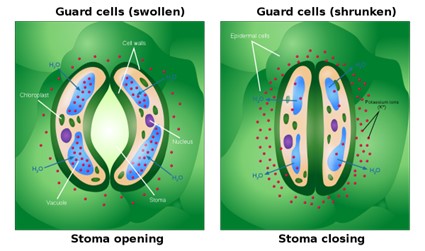
Figure 10: Opening and Closing of Stoma. As K+ levels increase in the guard cells, the water potential of the guard cells drops, and water enters the guard cells. Ali Zifan, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
Plants have evolved over time to adapt to their local environment and reduce transpiration. Desert plants (xerophytes) and plants that grow on other plants (epiphytes) have limited access to water. Such plants usually have a much thicker waxy cuticle than those growing in more moderate, well-watered environments (mesophytes). Aquatic plants (hydrophytes) also have their own set of anatomical and morphological leaf adaptations. Xerophytes and epiphytes often have a thick covering of trichomes or of stomata that are sunken below the leaf’s surface. Trichomes are specialized hair-like epidermal cells that secrete oils and substances. These adaptations impede air flow across the stomatal pore and reduce transpiration. Multiple epidermal layers are also commonly found in these types of plants.
Water & Solute Potential
Plants are phenomenal hydraulic engineers. Using only the basic laws of physics and the simple manipulation of potential energy, plants can move water to the top of a 116-meter-tall tree. Plants can also use hydraulics to generate enough force to split rocks and buckle sidewalks. Plants achieve this because of water potential.
Water potential is a measure of the potential energy in water. Plant physiologists are not interested in the energy in any one particular aqueous system, but are very interested in water movement between two systems. In practical terms, therefore, water potential is the difference in potential energy between a given water sample and pure water (at atmospheric pressure and ambient temperature). Water potential is denoted by the Greek letter ψ (psi) and is expressed in units of pressure (pressure is a form of energy) called megapascals (MPa). The potential of pure water (Ψwpure H2O) is, by convenience of definition, designated a value of zero (even though pure water contains plenty of potential energy, that energy is ignored). Water potential values for the water in a plant root, stem, or leaf are therefore expressed relative to Ψwpure H2O.
The water potential in plant solutions is influenced by solute concentration, pressure, gravity, and factors called matrix effects. Water potential can be broken down into its individual components using the following equation:
Ψsystem = Ψtotal = Ψs + Ψp + Ψg + Ψm
where Ψs, Ψp, Ψg, and Ψm refer to the solute, pressure, gravity, and matric potentials, respectively. “System” can refer to the water potential of the soil water (Ψsoil), root water (Ψroot), stem water (Ψstem), leaf water (Ψleaf) or the water in the atmosphere (Ψatmosphere): whichever aqueous system is under consideration. As the individual components change, they raise or lower the total water potential of a system. When this happens, water moves to equilibrate, moving from the system or compartment with a higher water potential to the system or compartment with a lower water potential. This brings the difference in water potential between the two systems (ΔΨ) back to zero (ΔΨ = 0). Therefore, for water to move through the plant from the soil to the air (a process called transpiration), Ψsoil must be > Ψroot > Ψstem > Ψleaf > Ψatmosphere.
Water only moves in response to ΔΨ, not in response to the individual components. However, because the individual components influence the total Ψsystem, by manipulating the individual components (especially Ψs), a plant can control water movement.
Solute potential (Ψs), also called osmotic potential, is negative in a plant cell and zero in distilled water. Typical values for cell cytoplasm are –0.5 to –1.0 MPa. Solutes reduce water potential (resulting in a negative Ψw) by consuming some of the potential energy available in the water. Solute molecules can dissolve in water because water molecules can bind to them via hydrogen bonds; a hydrophobic molecule like oil, which cannot bind to water, cannot go into solution. The energy in the hydrogen bonds between solute molecules and water is no longer available to do work in the system because it is tied up in the bond. In other words, the amount of available potential energy is reduced when solutes are added to an aqueous system. Thus, Ψs decreases with increasing solute concentration. Because Ψs is one of the four components of Ψsystem or Ψtotal, a decrease in Ψs will cause a decrease in Ψtotal. The internal water potential of a plant cell is more negative than pure water because of the cytoplasm’s high solute content (Fig 11). Because of this difference in water potential water will move from the soil into a plant’s root cells via the process of osmosis. This is why solute potential is sometimes called osmotic potential.
Plant cells can metabolically manipulate Ψs, and by extension, Ψtotal by adding or removing solute molecules. Therefore, plants have control over Ψtotal via their ability to exert metabolic control over Ψs.
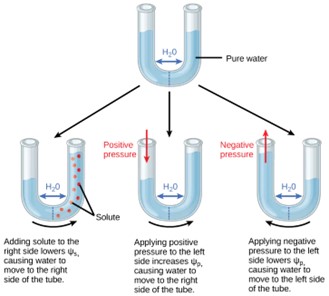
Figure 11: In this example with a semipermeable membrane between two aqueous systems, water will move from a region of higher to lower water potential until equilibrium is reached. Solutes (Ψs), pressure (Ψp), and gravity (Ψg) influence total water potential for each side of the tube (Ψtotal right or left), and therefore, the difference between Ψtotal on each side (ΔΨ). (Ψm , the potential due to interaction of water with solid substrates, is ignored in this example because glass is not especially hydrophilic). Water moves in response to the difference in water potential between two systems (the left and right sides of the tube).
Pressure potential (Ψp), also called turgor potential, may be positive or negative (Fig 12). Because pressure is an expression of energy, the higher the pressure, the more potential energy in a system, and vice versa. Therefore, a positive Ψp (compression) increases Ψtotal, and a negative Ψp (tension) decreases Ψtotal. Positive pressure inside cells is contained by the cell wall, producing turgor pressure. Pressure potentials are typically around 0.6–0.8 MPa, but can reach as high as 1.5 MPa in a well-watered plant. A Ψp of 1.5 MPa equates to 210 pounds per square inch (1.5 MPa x 140 lb in-2 MPa-1 = 210 lb/in-2). As a comparison, most automobile tires are kept at a pressure of 30–34 psi. An example of the effect of turgor pressure is the wilting of leaves and their restoration after the plant has been watered (Fig 12). Water is lost from the leaves via transpiration (approaching Ψp = 0 MPa at the wilting point) and restored by uptake via the roots.
A plant can manipulate Ψp via its ability to manipulate Ψs and by the process of osmosis. If a plant cell increases the cytoplasmic solute concentration, Ψs will decline, Ψtotal will decline, the ΔΨ between the cell and the surrounding tissue will decline, water will move into the cell by osmosis, and Ψp will increase. Ψp is also under indirect plant control via the opening and closing of stomata. Stomatal openings allow water to evaporate from the leaf, reducing Ψp and Ψtotal of the leaf and increasing ii between the water in the leaf and the petiole, thereby allowing water to flow from the petiole into the leaf.

Figure 12: When (a) total water potential (Ψtotal) is lower outside the cells than inside, water moves out of the cells and the plant wilts. When (b) the total water potential is higher outside the plant cells than inside, water moves into the cells, resulting in turgor pressure (Ψp) and keeping the plant erect. (credit: modification of work by Victor M. Vicente Selvas)
The following video does a nice job reminding you about osmosis and general concepts before reviewing water potential (starting around 7.25 minutes).
Transport of Sugars/Photosynthates in Plants
Plants need an energy source to grow. In seeds and bulbs, food is stored in polymers (such as starch) that are converted by metabolic processes into sucrose for newly developing plants. Once green shoots and leaves are growing, plants are able to produce their own food by photosynthesizing. The products of photosynthesis are called photosynthates, which are usually in the form of simple sugars such as sucrose.
Structures that produce photosynthates for the growing plant are referred to as sources. Sugars produced in sources, such as leaves, need to be delivered to growing parts of the plant via the phloem in a process called translocation. The points of sugar delivery, such as roots, young shoots, and developing seeds, are called sinks. Seeds, tubers, and bulbs can be either a source or a sink, depending on the plant’s stage of development and the season.
Movement of sugars from the cells into the phloem requires energy/ATP. This means active transport is needed. When the sugars arrive at the sinks, they will passively move into those cells. This means that no ATP energy is required. The dissolved sugars will diffuse from an area of high concentration (in the phloem) to an area of low concentration (in the sink cells).
The products from the source are usually translocated to the nearest sink through the phloem. For example, the highest leaves will send photosynthates upward to the growing shoot tip, whereas lower leaves will direct photosynthates downward to the roots. Intermediate leaves will send products in both directions, unlike the flow in the xylem, which is always unidirectional (soil to leaf to atmosphere). The pattern of photosynthate flow changes as the plant grows and develops. Photosynthates are directed primarily to the roots early on, to shoots and leaves during vegetative growth, and to seeds and fruits during reproductive development. They are also directed to tubers for storage.
So how does it work in more detail? Photosynthates, such as sucrose, are produced in the mesophyll cells of photosynthesizing leaves. Then ATP energy is used to actively move the excess sugars out of the leaf cells and into the phloem cells. When these sugars arrive, they lower the concentration of water in the phloem. As a result, water diffuses out of the xylem, which is adjacent to the phloem, and moves into the phloem. This osmosis is automatic and passive. That is, when the sugars are loaded into the phloem, water will move from a higher concentration in the xylem to a lower concentration in the phloem (in other words, osmosis occurs). The now dissolved sugars, called phloem sap, are found in a high turgor pressure environment. This fluid will now move toward a sink as a result (Fig 13b), where there is lower turgor pressure. Once at the sink, the sugars will diffuse out of the phloem and into the sink cells. Sucrose concentration in the sink cells is lower than in the phloem because the sink sucrose has been metabolized for growth, or converted to starch for storage or other polymers, such as cellulose, for structural integrity. Unloading at the sink end of the phloem tube occurs by either diffusion or active transport of sucrose molecules from an area of high concentration to one of low concentration. Water diffuses from the phloem by osmosis and is then transpired or recycled via the xylem back into the phloem sap.
So the phloem translocates sugars and larger molecules to where they are used or stored. The photosynthates in the leaf cells move through plasmodesmata channels (cytoplasmic channels) to reach specifically the phloem sieve-tube elements (STEs) in the vascular bundles. The sieve plates have pores that allow for pressure-driven bulk flow, or translocation, of phloem sap. Companion cells are associated with STEs. They assist with metabolic activities and produce energy for the STEs (Fig 13a). As the sucrose is actively transported against its concentration gradient into the phloem cells, an electrochemical potential of the proton gradient is used. This is coupled to the uptake of sucrose with a carrier protein called the sucrose-H+ symporter.
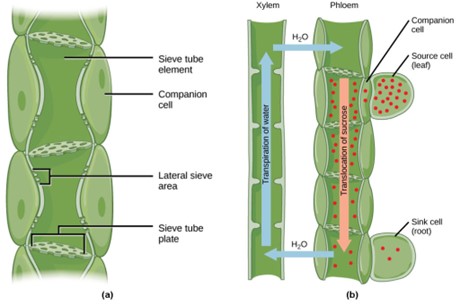
Figure 13: (a) Phloem is comprised of cells called sieve-tube elements. Phloem sap travels through perforations called sieve tube plates. Neighboring companion cells carry out metabolic functions for the sieve-tube elements and provide them with energy. Lateral sieve areas connect the sieve-tube elements to the companion cells. (b) Sucrose is actively transported from source cells into companion cells and then into the sieve-tube elements. This reduces the water potential, which causes water to enter the phloem from the xylem. The resulting positive pressure forces the sucrose-water mixture down toward the roots, where sucrose is unloaded. Transpiration causes water to return to the leaves through the xylem vessels.
This animation is a review of movement of food and sugars in the phloem.
Summary
Water movement in plants is essential for various physiological processes, including photosynthesis, nutrient uptake, and structural support. Water can only enter the root hairs of a plant and then it is pulled up through the xylem according to the transpiration-adhesion-cohension tension model (TACT). Here we find water ascending the plant’s stem and branches via capillary action and cohesion-adhesion forces. Capillary action occurs as water molecules stick to the interior walls of narrow xylem vessels and move upward against gravity due to their cohesion (attraction to each other) and adhesion (attraction to the vessel walls). The primary driving force for water movement in plants is transpiration, the loss of water vapor through small openings called stomata on the plant’s leaves. This loss creates a negative pressure gradient, known as tension, in the xylem. As water molecules evaporate from the leaves, they pull up water from the roots to replace the lost moisture.
The xylem vessels serve as the main conduits for water transport within the plant. These vessels consist of dead, hollow cells that provide a continuous pathway for water to move from the roots to the leaves and other plant parts.
Sugars, produced during photosynthesis in the leaves, need to be transported to various parts of the plant for energy production, growth, and storage. This process primarily involves the phloem, another specialized vascular tissue, and is known as translocation. Initially, sugar loading from a source occurs. After photosynthesis, sugars, primarily in the form of sucrose, are actively transported into the companion cells located near the phloem sieve tubes. Sugars are then transported through the phloem using a pressure flow mechanism. This mechanism relies on the creation of a pressure gradient between source and sink tissues. Source tissues, typically mature leaves, have a high concentration of sugars. Sink tissues, such as roots, developing leaves, and fruits, have a lower concentration. The sugars that are actively loaded into the sieve tube elements of phloem in the source tissues. This loading creates a high solute concentration in the sieve tubes, resulting in water uptake through osmosis. This influx of water creates positive pressure, known as turgor pressure, in the source tissue. The pressure gradient drives the movement of sap through the phloem tubes, effectively transporting sugars to where they are needed for growth, energy, and storage.
In summary, water moves through a plant via the xylem through a combination of root absorption, root pressure, capillary action, and transpiration, while sugars move through the phloem via active transport and pressure flow mechanisms to supply energy and nutrients to different parts of the plant. These two vascular systems play crucial roles in maintaining plant health and growth.
Questions
Glossary
References
Bellairs, S. Micrographs of Xylem and Phloem. Plant Anatomy and Physiology. Charles Darwin University.
Kosal, E. 2023. Summary. NC State University.
Lumen Learning. Content on Water and Solute Potential, and Plant Transport and Cells. Fundamentals of Biology.
