
14 The Seed Plants
By the end of this chapter you will be able to:
- Describe the distinguishing traits of the gymnosperms
- Describe the distinguishing traits of the angiosperms
- Describe the two major innovations that allowed seed plants to reproduce in the absence of water
- Explain when seed plants first appeared and when gymnosperms became the dominant plant group
- Discuss the purpose of pollen grains and seeds
- Describe the significance of angiosperms bearing both flowers and fruit
- List the four groups of modern-day gymnosperms and provide examples of each
- Describe the life cycle of a typical gymnosperm
- Explain why angiosperms are the dominant form of plant life in most terrestrial ecosystems
- Describe the main parts of a flower and their functions
- Discuss the similarities and differences between the two main groups of flowering plants
Introduction
In seed plants, the evolutionary trend led to a dominant sporophyte generation accompanied by a corresponding reduction in the size of the gametophyte from a conspicuous structure to a microscopic cluster of cells enclosed in the tissues of the sporophyte. Whereas lower vascular plants, such as club mosses and ferns, are mostly homosporous (producing only one type of spore), all seed plants, or spermatophytes, are heterosporous, producing two types of spores: megaspores (female) and microspores (male). Megaspores develop into female gametophytes that produce eggs, and microspores mature into male gametophytes that generate sperm. Because the gametophytes mature within the spores, they are not free-living, as are the gametophytes of other seedless vascular plants.
Ancestral heterosporous seedless plants, represented by modern-day plants such as the spike moss Selaginella, are seen as the evolutionary forerunners of seed plants. In the life cycle of Selaginella, both male and female sporangia develop within the same stem-like strobilus. In each male sporangium, multiple microspores are produced by meiosis. Each microspore produces a small antheridium contained within a spore case. As it develops it is released from the strobilus, and a number of flagellated sperm are produced that then leave the spore case. In the female sporangium, a single megaspore mother cell undergoes meiosis to produce four megaspores. Gametophytes develop within each megaspore, consisting of a mass of tissue that will later nourish the embryo and a few archegonia. The female gametophyte may remain within remnants of the spore wall in the megasporangium until after fertilization has occurred and the embryo begins to develop. This combination of an embryo and nutritional cells is a little different from the organization of a seed, since the nutritive endosperm in a seed is formed from a single cell rather than multiple cells.
Both seeds and pollen distinguish seed plants from seedless vascular plants. These innovative structures allowed seed plants to reduce or eliminate their dependence on water for gamete fertilization and development of the embryo, and to conquer dry land. Pollen grains are male gametophytes, which contain the sperm (gametes) of the plant. The small haploid (1n) cells are encased in a protective coat that prevents desiccation (drying out) and mechanical damage. Pollen grains can travel far from their original sporophyte, spreading the plant’s genes. Seeds offer the embryo protection, nourishment, and a mechanism to maintain dormancy for tens or even thousands of years, ensuring that germination can occur when growth conditions are optimal. Seeds therefore allow plants to disperse the next generation through both space and time. With such evolutionary advantages, seed plants have become the most successful and familiar group of plants.
Both adaptations expanded the colonization of land begun by the bryophytes and their ancestors. Fossils place the earliest distinct seed plants at about 350 million years ago. The first reliable record of gymnosperms dates their appearance to the Pennsylvanian period, about 319 million years ago (Figure 1). Gymnosperms were preceded by progymnosperms, the first naked seed plants, which arose about 380 million years ago. Progymnosperms were a transitional group of plants that superficially resembled conifers (cone bearers) because they produced wood from the secondary growth of the vascular tissues; however, they still reproduced like ferns, releasing spores into the environment. At least some species were heterosporous. Progymnosperms, like the extinct Archaeopteris (not to be confused with the ancient bird Archaeopteryx), dominated the forests of the late Devonian period (Fig 1). However, by the early (Triassic, c. 240 MYA) and middle (Jurassic, c. 205 MYA) Mesozoic era, the landscape was dominated by the true gymnosperms. Angiosperms surpassed gymnosperms by the middle of the Cretaceous (c. 100 MYA) in the late Mesozoic era, and today are the most abundant and biologically diverse plant group in most terrestrial biomes.

Figure 1: A 3D model of the tree-like plant Archaeopteris which lived in the Devonian period as was a precursor to the modern plant group, Gymnosperms. From Wikipedia Commons.
The modern vascular seed plants we find today are the Gymnosperms and the Angiosperms, the Flowering Plants. Notice on the timeline below (Fig 2) that Gymnosperms appeared around 330 milion years ago and the youngest plants, the flowering plants around 100 million years ago.
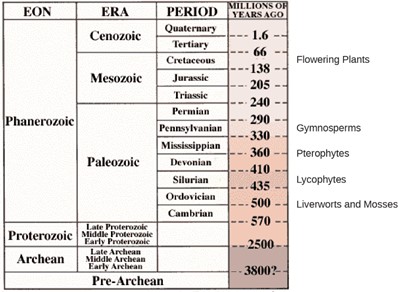
Figure 2: Plant timeline. Various plant species evolved in different eras. (credit: United States Geological Survey) Figure modified from source.
Bryophyte and fern spores are haploid cells dependent on moisture for rapid development of multicellular gametophytes. In the seed plants, the female gametophyte consists of just a few cells: the egg and some supportive cells, including the endosperm-producing cell that will support the growth of the embryo. After fertilization of the egg, the diploid zygote produces an embryo that will grow into the sporophyte when the seed germinates. Storage tissue to sustain growth of the embryo and a protective coat give seeds their superior evolutionary advantage. Several layers of hardened tissue prevent desiccation, and free the embryo from the need for a constant supply of water. Furthermore, seeds remain in a state of dormancy—induced by desiccation and the hormone abscisic acid—until conditions for growth become favorable. Whether blown by the wind, floating on water, or carried away by animals, seeds are scattered in an expanding geographic range, thus avoiding competition with the parent plant.
Pollen grains (Fig 3) are male gametophytes containing just a few cells and are distributed by wind, water, or an animal pollinator. The whole structure is protected from desiccation and can reach the female organs without depending on water. After reaching a female gametophyte, the pollen grain grows a tube that will deliver a male nucleus to the egg cell. The sperm of modern gymnosperms and all angiosperms lack flagella, but in cycads, Ginkgo, and other primitive gymnosperms, the sperm are still motile, and use flagella to swim to the female gamete; however, they are delivered to the female gametophyte enclosed in a pollen grain. The pollen grows or is taken into a fertilization chamber, where the motile sperm are released and swim a short distance to an egg.
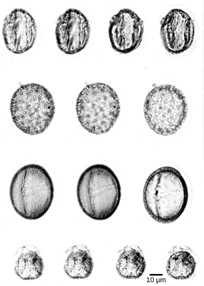
Figure 3: Pollen fossils. This fossilized pollen is from a Buckbean fen core found in Yellowstone National Park, Wyoming. The pollen is magnified 1,054 times. (credit: R.G. Baker, USGS; scale-bar data from Matt Russell)
The roughly 200 million years between the appearance of the gymnosperms and the flowering plants gives us some appreciation for the evolutionary experimentation that ultimately produced flowers and fruit. Angiosperms (“seed in a vessel”) produce a flower containing male and/or female reproductive structures. Fossil evidence indicates that flowering plants first appeared about 125 million years ago in the Lower Cretaceous (late in the Mesozoic era), and were rapidly diversifying by about 100 million years ago in the Middle Cretaceous. Earlier traces of angiosperms are scarce. Fossilized pollen recovered from Jurassic geological material has been attributed to angiosperms. A few early Cretaceous rocks show clear imprints of leaves resembling angiosperm leaves. By the mid-Cretaceous, a staggering number of diverse flowering plants crowd the fossil record. The same geological period is also marked by the appearance of many modern groups of insects, suggesting that pollinating insects played a key role in the evolution of flowering plants.
New data in comparative genomics and paleobotany (the study of ancient plants) have shed some light on the evolution of angiosperms. Although the angiosperms appeared after the gymnosperms, they are probably not derived from gymnosperm ancestors. Instead, the angiosperms form a sister clade (a species and its descendants) that developed in parallel with the gymnosperms. The two innovative structures of flowers and fruit represent an improved reproductive strategy that served to protect the embryo while increasing genetic variability and range. There is no current consensus on the origin of the angiosperms. Paleobotanists debate whether angiosperms evolved from small woody bushes or were related to the ancestors of tropical grasses. Both views draw support from cladistics, and the so-called woody magnoliid hypothesis—which proposes that the early ancestors of angiosperms were shrubs like modern magnolia—also offers molecular biological evidence.
The most primitive living angiosperm is considered to be Amborella trichopoda, a small plant native to the rainforest of New Caledonia, an island in the South Pacific (Fig 4). Analysis of the genome of A. trichopoda has shown that it is related to all existing flowering plants and belongs to the oldest confirmed branch of the angiosperm family tree. The nuclear genome shows evidence of an ancient whole-genome duplication. The mitochondrial genome is large and multichromosomal, containing elements from the mitochondrial genomes of several other species, including algae and a moss. A few other angiosperm groups, called basal angiosperms, are viewed as having ancestral traits because they branched off early from the phylogenetic tree. Most modern angiosperms are classified as either monocots or eudicots, based on the structure of their leaves and embryos (Fig 5). Basal angiosperms, such as water lilies, are considered more ancestral in nature because they share morphological traits with both monocots and eudicots.

Figure 4: Amborella trichopoda showing male flowers. Scott Zona from Miami, Florida, CC via Wikimedia Commons
The most easy to distinguish traits when looking at whether an angiosperm is a monocot or a dicot has to do with the flowers. If you see floral parts in multiples of 3, it is a monocot and if you see multiples of 4 or 5, it is a dicot (Fig 5). The veins can also give a hint, as monocots have parallel veins and dicots a network of veins (Fig 5).

Figure 5: This diagram is showing the differences between monocotyledonous plants and dicotyledonous plants. Monocots have a single cotyledon and long and narrow leaves with parallel veins. Their vascular bundles are scattered. Their petals or flower parts are in multiples of three. Dicots have two cotyledons and broad leaves with network of veins. Their vascular bundles are in a ring. Their petals or flower parts are in multiples of four or five. Flowerpower207, CC via Wikimedia Commons
Angiosperms produce their gametes in separate organs, which are usually housed in a flower. Both fertilization and embryo development take place inside an anatomical structure that provides a stable system of sexual reproduction largely sheltered from environmental fluctuations. With about 300,000 species, flowering plants are the most diverse phylum on Earth after insects, which number about 1,200,000 species. Flowers come in a bewildering array of sizes, shapes, colors, smells, and arrangements. Most flowers have a mutualistic pollinator, with the distinctive features of flowers reflecting the nature of the pollination agent. The relationship between pollinator and flower characteristics is one of the great examples of coevolution.
Following fertilization of the egg, the ovule grows into a seed. The surrounding tissues of the ovary thicken, developing into a fruit that will protect the seed and often ensure its dispersal over a wide geographic range. Not all fruits develop completely from an ovary; such “false fruits” or pseudocarps, develop from tissues adjacent to the ovary. Like flowers, fruit can vary tremendously in appearance, size, smell, and taste. Tomatoes, green peppers, corn, and avocados are all examples of fruits. Along with pollen and seeds, fruits also act as agents of dispersal. Some may be carried away by the wind. Many attract animals that will eat the fruit and pass the seeds through their digestive systems, then deposit the seeds in another location. Cockleburs are covered with stiff, hooked spines that can hook into fur (or clothing) and hitch a ride on an animal for long distances. The cockleburs that clung to the velvet trousers of an enterprising Swiss hiker, George de Mestral, inspired his invention of the loop and hook fastener he named Velcro.

Gymnosperms
Gymnosperms, meaning “naked seeds,” are a diverse group of seed plants. According to the “anthophyte” hypothesis, the angiosperms are a sister group of one group of gymnosperms (the Gnetales), which makes the gymnosperms a paraphyletic group. Paraphyletic groups are those in which not all descendants of a single common ancestor are included in the group. However , the “netifer” hypothesis suggests that the gnetophytes are sister to the conifers, making the gymnosperms monophyletic and sister to the angiosperms. Further molecular and anatomical studies may clarify these relationships. Characteristics of the gymnosperms include naked seeds, separate female and male gametophytes, pollen cones and ovulate cones, pollination by wind and insects, and tracheids (which transport water and solutes in the vascular system).
Gymnosperm seeds are not enclosed in an ovary; rather, they are only partially sheltered by modified leaves called sporophylls. You may recall the term strobilus (plural = strobili) describes a tight arrangement of sporophylls around a central stalk, as seen in pine cones. Some seeds are enveloped by sporophyte tissues upon maturation. The layer of sporophyte tissue that surrounds the megasporangium, and later, the embryo, is called the integument. The “typical” gymnosperm will contain small male pine cones, where pollen is produced, and larger female pine cones, where the egg is found. The pollen will land on the larger female cone and need to grow a pollen tube to eventually reach the egg. This process can take years. Once the sperm nucleus moves through the pollen tube, it can fertilize the egg nucleus to form an embryo in the seed (Fig 6).
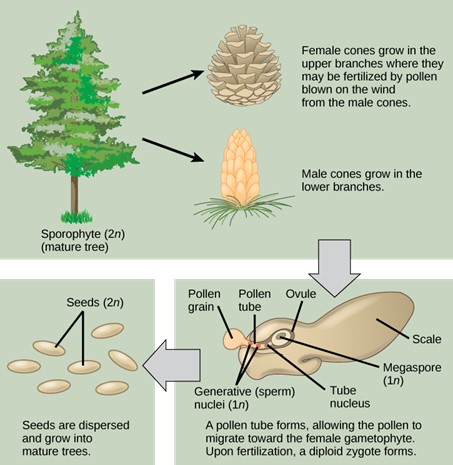
Figure 6: The conifer life cycle. This image shows the life cycle of a conifer. Pollen from male cones blows up into upper branches, where it fertilizes the eggs in female cones. The megspore shown in the image develops into the female gametophyte as the pollen tube slowly grows toward it, eventually fusing with the egg and delivering a male sperm nucleus, which combines with the female nucleus of the mature egg to form an embryo.
Watch this video on the life cycle of gymnosperms.
Gymnosperms were the dominant phylum in the Mesozoic era. They are adapted to live where fresh water is scarce during part of the year, or in the nitrogen-poor soil of a bog. Therefore, they are still the prominent phylum in the coniferous biome or taiga, where the evergreen conifers have a selective advantage in cold and dry weather. Evergreen conifers continue low levels of photosynthesis during the cold months, and are ready to take advantage of the first sunny days of spring. One disadvantage is that conifers are more susceptible than deciduous trees to leaf infestations because most conifers do not lose their leaves all at once. They cannot, therefore, shed parasites and restart with a fresh supply of leaves in spring.
Although most gymnosperms have needles, some do have leaves that are deciduous. Examples include the Ginkgo tree (Fig 7). And although most gymnosperms have cones, some species do have different structures like the Ginkgo (Fig 7).

Figure 7: The Ginkgo tree is a gymnosperm even though it does not bear needles or cones. The Ginkgo is deciduous and can lose its leaves in the autumn. Jalexlb, CC, via Wikimedia Commons
The life cycle of a gymnosperm involves alternation of generations, with a dominant sporophyte in which reduced male and female gametophytes reside. All gymnosperms are heterosporous. The male and female reproductive organs can form in cones or strobili. Male and female sporangia are produced either on the same plant, described as monoecious (“one home” or bisexual), or on separate plants, referred to as dioecious (“two homes” or unisexual) plants. The life cycle of a conifer will serve as our example of reproduction in gymnosperms.
Modern gymnosperms are classified into four phyla. Coniferophyta, Cycadophyta, and Ginkgophyta are similar in their pattern of seed development and also in their production of secondary cambium (cells that generate the vascular system of the trunk or stem and are partially specialized for water transportation). However, the three phyla are not closely related phylogenetically to each other. Gnetophyta are considered the closest group to angiosperms because they produce true xylem tissue, with vessels as well as the tracheids found in the rest of the gymnosperms. It is possible that vessel elements arose independently in the two groups.
Conifers (Coniferophyta)
Conifers are the dominant phylum of gymnosperms, with the greatest variety of species (Fig 8). Typical conifers are tall trees that bear scale-like or needle-like leaves. Water evaporation from leaves is reduced by their narrow shape and a thick cuticle. Snow easily slides off needle-shaped leaves, keeping the snow load light, thus reducing broken branches. Such adaptations to cold and dry weather explain the predominance of conifers at high altitudes and in cold climates. Conifers include familiar evergreen trees such as pines, spruces, firs, cedars, sequoias, and yews. A few species are deciduous and lose their leaves in fall. The bald cypress, dawn redwood, European larch and the tamarack (Fig 8) are examples of deciduous conifers. Many coniferous trees are harvested for paper pulp and timber. The wood of conifers is more primitive than the wood of angiosperms; it contains tracheids, but no vessel elements, and is therefore referred to as “soft wood.”

Figure 8: Conifers. Conifers are the dominant form of vegetation in cold or arid environments and at high altitudes. Shown here are the (a) evergreen spruce Picea sp., (b) juniper Juniperus sp., (c) coastal redwood or sequoia Sequoia sempervirens, and (d) the tamarack Larix laricina. Notice the deciduous yellow leaves of the tamarack. (credit a: modification of work by Rosendahl; credit b: modification of work by Alan Levine; credit c: modification of work by Wendy McCormic; credit d: modification of work by Micky Zlimen)
Cycads
Cycads thrive in mild climates and are often mistaken for palms because of the shape of their large, compound leaves. Cycads bear large strobili or cones (Fig 9), and may be pollinated by beetles rather than wind, which is unusual for a gymnosperm. Large cycads dominated the landscape during the age of dinosaurs in the Mesozoic, but only a hundred or so smaller species persisted to modern times. They face possible extinction, and several species are protected through international conventions. Because of their attractive shape, they are often used as ornamental plants in gardens in the tropics and subtropics.

Figure 9: This cycad, Encephalartos ferox, has large cones and broad, fern-like leaves. (credit: Wendy Cutler)
Ginkgophytes
The single surviving species of the ginkgophytes group is Ginkgo biloba mentioned earlier (Fig 7). Its fan-shaped leaves—unique among seed plants because they feature a dichotomous venation pattern—turn yellow in autumn and fall from the tree. For centuries, G. biloba was cultivated by Chinese Buddhist monks in monasteries, which ensured its preservation. It is planted in public spaces because it is unusually resistant to pollution. Male and female organs are produced on separate plants. Typically, gardeners plant only male trees because the seeds produced by the female plant have an off-putting smell of rancid butter.
Gnetophytes
The phylogenetic position of the gnetophytes is not currently resolved. Their possession of vessel elements suggests they are the closest relative to modern angiosperms. However, molecular analysis places them closer to the conifers. The three living genera are quite dissimilar: Ephedra, Gnetum, and Welwitschia (Fig 10), which may indicate that the group is not monophyletic. Like angiosperms, they have broad leaves. Ephedra (Fig 10a) occurs in dry areas of the West Coast of the United States and Mexico. Ephedra’s small, scale-like leaves are the source of the compound ephedrine, which is used in medicine as a potent decongestant. Because ephedrine is similar to amphetamines, both in chemical structure and neurological effects, its use is restricted to prescription drugs. Gnetum species (Fig 10b) are found in some parts of Africa, South America, and Southeast Asia, and include trees, shrubs and vines. Welwitschia (Fig 10c) is found in the Namib desert, and is possibly the oddest member of the group. It produces only two leaves, which grow continuously throughout the life of the plant (some plants are hundreds of years old). Like the ginkgos, Welwitschia produces male and female gametes on separate plants.

Figure 10: (a) Ephedra viridis, known by the common name Mormon tea, grows on the West Coast of the United States and Mexico. (b) Gnetum gnemon grows in Malaysia. (c) The large Welwitschia mirabilis can be found in the Namibian desert. (credit a: modification of work by USDA; credit b: modification of work by Malcolm Manners; credit c: modification of work by Derek Keats)
Watch this short video on Welwitschia plants – so interesting!
Angiosperms
From their humble and still obscure beginning during the early Jurassic period, the angiosperms—or flowering plants—have evolved to dominate most terrestrial ecosystems. With more than 300,000 species, the angiosperm phylum (Anthophyta) is second only to insects in terms of diversification.
The success of angiosperms is due to two novel reproductive structures: flowers and fruits. The function of the flower is to ensure pollination, often by arthropods, as well as to protect a developing embryo. The colors and patterns on flowers offer specific signals to many pollinating insects or birds and bats that have coevolved with them. For example, some patterns are visible only in the ultraviolet range of light, which can be seen by arthropod pollinators. For some pollinators, flowers advertise themselves as a reliable source of nectar. Flower scent also helps to select its pollinators. Sweet scents tend to attract bees and butterflies and moths, but some flies and beetles might prefer scents that signal fermentation or putrefaction. Flowers also provide protection for the ovule and developing embryo inside a receptacle. The function of the fruit is seed protection and dispersal. Different fruit structures or tissues on fruit—such as sweet flesh, wings, parachutes, or spines that grab—reflect the dispersal strategies that help spread seeds.
Flowers are modified leaves, or sporophylls, organized around a central receptacle (Fig 11). Although they vary greatly in appearance, virtually all flowers contain the same structures: sepals, petals, carpels, and stamens. The peduncle typically attaches the flower to the plant proper. A whorl of sepals (collectively called the calyx) is located at the base of the peduncle and encloses the unopened floral bud. Sepals are usually photosynthetic organs, although there are some exceptions. For example, the corolla in lilies and tulips consists of three sepals and three petals that look virtually identical. Petals, collectively the corolla, are located inside the whorl of sepals and may display vivid colors to attract pollinators. Sepals and petals together form the perianth. The sexual organs, the female gynoecium and male androecium are located at the center of the flower. Typically, the sepals, petals, and stamens are attached to the receptacle at the base of the gynoecium, but the gynoecium may also be located deeper in the receptacle, with the other floral structures attached above it.
The innermost part of a perfect flower is the gynoecium, the location in the flower where the eggs will form. The female reproductive unit consists of one or more carpels, each of which has a stigma, style, and ovary. The stigma is the location where the pollen is deposited either by wind or a pollinating arthropod. The sticky surface of the stigma traps pollen grains, and the style is a connecting structure through which the pollen tube will grow to reach the ovary. The ovary houses one or more ovules, each of which will ultimately develop into a seed. Flower structure is very diverse, and carpels may be singular, multiple, or fused. (Multiple fused carpels comprise a pistil.) The androecium, or male reproductive region is composed of multiple stamens surrounding the central carpel. Stamens are composed of a thin stalk called a filament and a sac-like structure called the anther. The filament supports the anther, where the microspores are produced by meiosis and develop into haploid pollen grains, or male gametophytes.
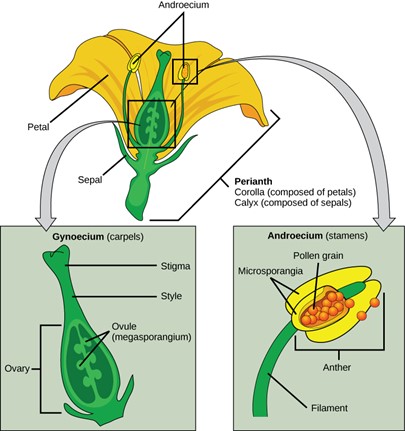
Figure 11: The structure of a perfect flower, with both male and female foral organs. The flower hown ha sonly one carpel, but some flowers have a cluster of carpels. Together, all the carpels make up the gynoecium Credit: modification of work by Mariana Ruiz Villareal)
What is unique to angiosperms is an event called double fertilization. This is when one sperm and the egg combine, forming a diploid zygote—the future embryo. The other sperm fuses with the polar nuclei, forming a triploid cell that will develop into the endosperm—the tissue that serves as a food reserve for the developing embryo. The zygote develops into an embryo with a radicle, or small root, and one (monocot) or two (dicot) leaf-like organs called cotyledons. This difference in the number of embryonic leaves is the basis for the two major groups of angiosperms: the monocots and the eudicots (Fig 4). Seed food reserves are stored outside the embryo, in the form of complex carbohydrates, lipids, or proteins. The cotyledons serve as conduits to transmit the broken-down food reserves from their storage site inside the seed to the developing embryo. The seed consists of a toughened layer of integuments forming the coat, the endosperm with food reserves, and at the center, the well-protected embryo.
As the seed develops, the walls of the ovary thicken and form the fruit. The seed forms in an ovary, which also enlarges as the seeds grow. Many foods commonly called vegetables are actually fruits. Eggplants, zucchini, string beans, tomatoes, and bell peppers are all technically fruits because they contain seeds and are derived from the thick ovary tissue. Acorns are true nuts, and winged maple “helicopter seeds” or whirligigs (whose botanical name is samara) are also fruits. Botanists classify fruit into more than two dozen different categories, only a few of which are actually fleshy and sweet.
Mature fruit can be fleshy or dry. Fleshy fruit include the familiar berries, peaches, apples, grapes, and tomatoes. Rice, wheat, and nuts are examples of dry fruit. Another subtle distinction is that not all fruits are derived from just the ovary. For instance, strawberries are derived from the ovary as well as the receptacle, and apples are formed from the ovary and the pericarp, or hypanthium. Some fruits are derived from separate ovaries in a single flower, such as the raspberry. Other fruits, such as the pineapple, form from clusters of flowers. Additionally, some fruits, like watermelon and orange, have rinds. Regardless of how they are formed, fruits are an agent of seed dispersal. The variety of shapes and characteristics reflect the mode of dispersal. Wind carries the light dry fruits of trees and dandelions. Water transports floating coconuts. Some fruits attract herbivores with their color or scent, or as food. Once eaten, tough, undigested seeds are dispersed through the herbivore’s feces (endozoochory). Other fruits have burrs and hooks to cling to fur and hitch rides on animals (epizoochory).
Summary
Seed plants are a diverse group of plants that have evolved specialized structures to produce and protect their seeds, allowing them to thrive in a wide range of environments. The two main categories of seed plants are gymnosperms and angiosperms, each with distinct characteristics and reproductive strategies. Gymnosperms are a group of seed plants that bear “naked” seeds, meaning that the seeds are not enclosed within a protective fruit. They were the dominant plants during the Mesozoic era and include several well-known plant types such as the conifers with their needles and cones, the cycads that are palm-like in appearance, the fan-shaped leafed ginkgos and the gnetophytes which are very diverse in form and have features that resemble not only the gymnosperms but also the angiosperms. The angiosperms are are the most widespread and diverse group of plants on Earth. They are characterized by producing seeds enclosed within a protective structure called a fruit. Angiosperms can be further classified into two main groups known as monocots and dicots, based on the number of cotyledons each group has, among other traits. Angiosperms have a remarkable variety of reproductive strategies, including insect and animal pollination, wind dispersal, and fruit consumption by animals for seed dispersal. Their adaptability and capacity for coevolution with pollinators and seed dispersers have contributed to their success in colonizing diverse habitats.
Angiosperms are most notable for their unique double fertilization. It involves the fertilization of two distinct structures within the ovule, resulting in the formation of both the embryo and endosperm, which nourishes the developing embryo. Double fertilization is a significant evolutionary innovation of angiosperms and plays a crucial role in their reproductive success. It ensures that the embryo has a nutrient-rich food source (endosperm) to sustain its early growth, enhancing the chances of successful germination and establishment of the new plant. This process also contributes to the efficient allocation of resources, allowing angiosperms to thrive in diverse ecological niches and environments.
Questions
Glossary
References
Clark, MA, Choi, J, and M. Douglas. Biology 2e for Biol 111 and Biol 112.
Kosal, E. 2023. Summary. NC State University.
