
7 Evidence of Evolution
At the end of this chapter, you will be able to:
- Explain the pieces of evidence to support evolution
- Distinguish between the different types of fossils
- Compare and contrast homologous structures to analogous structures
- Describe the role that hox genes play in the development of embryos
Introduction
Evolution is the change in the allelic frequency of a trait over time. Sometimes the changes are small resulting in the change of the variation of a trait in the same species – we would call this microevolution. Other times, so much DNA changes that we say a new species has formed, that is macroevolution has occurred. Regardless, there are different types of evidence that scientists use to support evolution. So much evidence has been collected from different disciplines, such as geology, embryology, genetics, and cell biology, that we refer to evolution as a theory. It has been supported by so many disciplines and has stood the test of time.
Physical Evidence
Fossils are the preserved remains or traces of organisms that lived in the remote past (over 10,000 years ago). There are many different types of fossils that occur in various steps of completeness, collectively forming the fossil record, which provides a wealth of information about evolution (Fig 1).

Figure 1: Examples of fossilized remains a) petrified wood from Petrified Forest National Park, Arizona; b) a fossilized flatfish; c) two 125-million-year-old dung midges, perfectly preserved in amber, are the oldest preserved mating pair in the animal kingdom; and d) fossilized dinosaur footprints.
Some fossils are mineralized remains that result from a process in which living tissues are replaced by minerals such as quartz, calcite, and silica. In some cases, a near-perfect representative of an organism is captured in stone—this is called petrifaction. Petrified wood looks a lot like a tree trunk or branch, but if you touch it or kick it, you’ll find out quickly that it’s made of stone. We have learned a lot from petrified wood; for example, the discovery of fossilized tree trunks and leaves in Antarctica suggests the icy landscape was once covered by lush green forests.
The remains of some organisms are only evident as casts or molds. Molds are impressions left by organisms, whereas casts represent molds that have been filled in by mineralization. In particular, sedimentary rock, which is formed by years of sand accumulating in layers on ancient sea floors, is an excellent source of fossil remains of aquatic organisms. For example, recent discoveries of fish casts have clarified some details about flatfish evolution. Flatfish, such as flounders, have eyes on one side of their heads, posing the question: how did such a body plan evolve? In the past several years, many examples of ancient fish with “transitional” eye arrangements have been discovered; specifically, these fish had eyes that were arranged intermediate to those of flounders and typical bony fish.
Some organisms persist long after death as unaltered remains. Unaltered remains have changed little since they died, and include hard, decay-resistant tissues such as teeth or shells. Soft tissues can also persist for thousands of years if they have been protected from heat or oxygen (both of which cause decay). Unaltered remains are typically found in ice, amber, or tar deposits. In 2012, an international team of scientists discovered two new species of small insects perfectly preserved in amber (fossilized tree resin) from over 100 million years ago. The discovery of these insects, called thrips, is exciting by itself, but the thrips also carried tiny pollen grains on their wings and abdomens. This is the oldest known evidence of pollination, whereby an animal carries a plant’s sperm (as pollen grains) to eggs. This is also the first example of pollination from the era of the dinosaurs.
Trace remains are fossilized evidence of living organisms, but not the actual organisms themselves. Examples of trace remains include footprints preserved in rock, mineralized feces (or coprolites), and the molds of once-inhabited burrows. The recent discovery of reptile footprints, in a location that would have been hundreds of miles from the sea, suggests that reptiles have been able to move into continental interiors for at least 318 million years. This discovery pushed back earlier estimates for the movement of reptiles away from the sea by several million years.
Related organisms have similar bodily structures, or anatomy; for example, the skeletons of humans, rats, and bats are similar, despite numerous obvious differences. In fact, there is a bone-by-bone similarity in the bodies of these three animals. Anatomical similarities are particularly evident in a comparison of the forelimbs of the pterodactyl, bat, hawk, whale, and human (Fig 2). These animals live in different types of environments, and use their limbs for different functions–flying, swimming, or grasping and throwing. Despite these different functions, all of these animals possess limbs with certain shared characteristics: a five-digit structure, a single large bone (humerus), two bones in the forearm (ulna and radius), and several wrist bones. These bones are homologous to each other, that is, they were inherited from a common ancestor.

Figure 2: Homologies in vertebrate forelimbs.
In fact, all four-limbed animals (or tetrapods)–amphibians, reptiles, birds, and mammals–have similar limbs. Anatomical similarities, such as those that characterize tetrapod limbs, are used as evidence for common ancestry. In other words, these similarities suggest that pterodactyls, bats, hawks, whales, and humans all share a common ancestor somewhere in their distant past. Learning about shared ancestry provides an important clue to understanding an organism’s evolution.
Homologous structures are anatomical features that different organisms share as a result of a common ancestor. The tetrapod limbs discussed above are homologous structures, or homologies. Because they evolved in an ancestor and are currently shared by different organisms, homologies are often called shared derived traits. Cactus spines, the leaves of a maple tree, and the cup-like “pitcher” of a pitcher plant are all modified from a common structure in an ancestor shared by all leaf-bearing land plants. While these leaves look different, and have evolved to serve different purposes, they are homologous structures and tell important stories about each plant’s history (Fig 3).

Figure 3: Leaves as homologous structures. Cactus spines, the leaves of a maple tree, and the cup-like “pitcher” of a pitcher plant are all modified from a common structure in an ancestor shared by all leaf-bearing land plants.
Analogous structures are anatomical features that different types of organisms share, but not as a result of a shared ancestor. Instead, analogous structures, or analogies, are similar because the organisms have changed in response to similar environments. When distantly related organisms share features as a result of similar environmental pressures (and not because of common ancestry), we say that they have undergone convergent evolution. For example, arctic mammals such as foxes and snowshoe hares grow white fur during the winter months. White fur allows these organisms to blend into the ice and snow that characterizes their polar home, and presumably protects them from predation. However, foxes and snowshoe hares do not share a common ancestor with white fur. Of course they ultimately share a common ancestor, as do all mammals, but the fox lineage is full of non-white animals, as is the group to which hares belong. The winter white of arctic foxes and snowshoe hares is thus an analogous structure, due to convergent evolution in a white, wintry landscape.
Vestigial structures are anatomical features that are either no longer in use, or their use has been greatly reduced or altered. Vestigial structures, or vestigia, provide clues to an organism’s history by suggesting what features were useful in the past, and by linking an organism to other, related organisms. For example, many mammals exhibit piloerection, whereby muscles constrict around the hair follicles and the animal’s hair stands on end. If you’ve ever surprised or otherwise threatened a dog or cat, you’ve probably seen the results of piloerection. Humans are mammals too, but we’ve been reduced to a scant covering of body hair. When we are scared, we also constrict the muscles around our hair follicles, but this just gives us goosebumps (Fig 4). Goosebumps aren’t very scary, but as vestigia they link us to our mammalian relatives, and tell us something about our evolutionary history.

Figure 4: The Vestigial piloerection of humans leads to goosebumps
Biogeographical Evidence
Biogeography, the study of the distribution of living organisms, addresses several evolutionary questions: How many types of organisms exist? Why are some types of organisms (e.g. insects) more abundant than others (e.g. mammals)? Why do certain organisms live in some places and not others? Why do islands have such distinct biodiversity compared to the larger continents? Why aren’t there any polar bears in Antarctica? Why aren’t there any giraffes in Hawaii? And so on….
Likewise, an understanding of evolution helps us appreciate the otherwise-perplexing global distribution of marsupial mammals (Fig 5). Marsupials are mammals in which females transport their young in distinctive pouches throughout their early infancy. In contrast, placental mammals have placental gestation and young that are born at a more developed stage. Marsupial mammals, such as kangaroos, opossums, wombats, and wallabies, occur in North and South America, Australia, and New Guinea, whereas the more-numerous placental mammals dominate the rest of the world. How can we explain these odd distribution patterns?
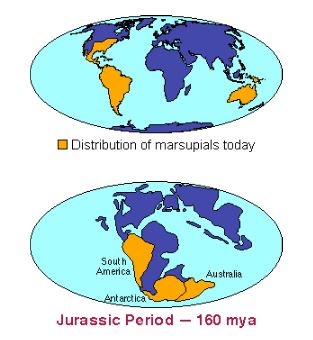
FIgure 5: Distribution of marsupials today versus Jurassic period
Knowledge of Earth’s history is key to biogeography and evolutionary understanding. Indeed, a consideration of continental drift—the movement of the continents, over geologic time, as a result of the movement of plates in Earth’s crust—is necessary to understand marsupial biogeography (Fig 6). We also know, from fossil evidence, that marsupials originated over 150 million years ago in China, at a time when the Asian and North American landmasses were joined. Marsupials dispersed to South America, and from there to Antarctica and Australia, both of which were attached to South America. When Australia, Antarctica, and South America drifted apart, each landmass carried a population of marsupial mammals. Over time, Australia’s marsupials evolved into the bandicoots, kangaroos, koalas, and other marsupials that inhabit the continent today. When Australia moved closer to Asia, about 15 million years ago, placental mammals such as rats and bats colonized the landmass.
Figure 7: Marsupial mammals
Genetic Evidence
DNA can provide a lot of information to help us study relationships among organisms. We can look at genes from 2 species to determine if they are similar or different to help us understand how closely or distantly related they are to one another. Homeotic genes are sections of DNA that play a key role in the developmental process as embryos are undergoing all sorts of changes. Homeotic genes act as on and off switches to help with the positioning of cells within the organism’s body plan.
One group of homeotic genes that turn out to be very helpful for understanding evolution are hox genes. Hox genes determine the form, number and evolution of repeating parts, such as the number and type of vertebrae in animals. The role of hox genes is to specify positional identity in the embryo which will lead to corresponding structures in the adult. At least eight different Hox gene are critical for the development of different body segment in fruti flies (Drosophila; Fig 8). Hox genes also affect th orientation of segments such that the anterior-to-posterior orientation is maintained. Scientists have learned that hox genes are part of a “toolbox” of genes that can be shuffled and rearranged in order to create the diversity of species we find on the planet.
When studying animal species, those animals with more simple body structures do not have as many hox genes as those species with more complex bodies. For example, a mammal such as a mouse will have many more hox genes than a flatworm. Humans have over 200 homeotic genes, and out of that, 39 are hox genes.
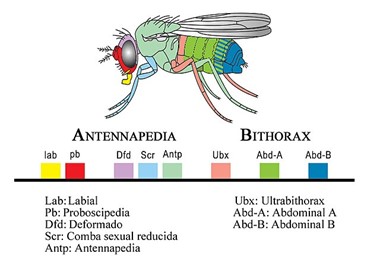
Figure 8: The hox genes arranged on a chromosome found in the fruit fly. Each box of color represents a gene, a section of DNA, that helps determine the body part on the fly highlighted in the same color. For example, the yellow hox gene “lab” will help to dictate the labial mouth part of the adult fly. Antonio Quesada Díaz, Public domain, via Wikimedia Commons
Plant species also have homeotic genes that can determine reproductive structures, such as stamens, carpel, and petals of flowering plants (Fig 9). Because flowers are the reproductive organs of angiosperms, a small change to the homeotic genes underlying the development of these strucurs can have a large impact on the phenotype and reproductive success of an individual and help drive evolution.

Figure 9: Parts of a Flower. The pistil comprises the female part and the stamen, the male part. LadyofHats, Public domain, via Wikimedia Commons
When looking more closely at homeotic genes, scientists discovered a 180-base-pair sequence to be identical in a wide array of animal species. This 180 base-pair sequence is called the homeobox and is highly conserved across evolutionary time (and it was first discovered in Drosophilia). This suggest that hox genes arose very early in evolutionary time. The shared hoemobox sequence suggest an ancestral gene was present and duplicated multiple times over evolutionary time to give rise to what we find today. Scientists get very excited about the homeobox because mutations in this gene can cause dramatic developmental changes in body parts and can lead to speciation.
Hox genes, as mentioned above, are also conserved across species. We find some of the human hox genes are homologous to those in the fruit fly. In experiments with mice, the Hox10 genes turn the “rib” genes off which are normally active in the lower back, but not needed. When scientists experimentally manipulated these genes they can cause ribs to grow in the vertebrae of the lower back. As such, we can see hox genes dictating arms versus legs as well as differentiating specific fingers, like a thumb or a pinky. The variation on the theme of the hox genes will give an organism a variety of traits and can lead to speciation.
Watch this animation on how these master genes control basic body plans: Fruit fly gene expression, development :: DNA from the Beginning (dnaftb.org)
Questions
Glossary
References
Cotner S. and D Wassenberg. The Evolution and Biology of Sex.
Genetic Science Learning Center. University of Utah. Homeotic Genes and Body Patterns. Found at https://learn.genetics.utah.edu/content/basics/hoxgenes/
Kosal, E. 2023. Genetics Evidence section. NC State University
