
4 Evolutionary Change Outside of Natural Selection
By the end of this chapter you will be able to:
- Explain why small populations undergo evolutionary change more rapidly than large populations
- Explain the role that the founder’s effect can have on evolution
- Compare and contrast genetic drift to natural selection
- Compare and contrast point and non-point mutations
Introduction
Natural selection is powerful as we learned in the previous chapter. Natural selection drives adaptive evolutionary change. Natural selection is not the only way, however, in which we can see populations evolve. It is the only way in which populations evolve adaptability.
We can see more random changes in a population through other evolutionary mechanisms, such as genetic drift, gene flow through immigration and emigration, and random mutations.
Genetic Drift
Genetic drift refers to changes in allele frequencies across generations that occur purely by random chance. The term “random” is key to an understanding of drift. Sometimes chance events, such as a natural disaster, influence which individuals survive and reproduce and thus influence the allele frequencies in the next generation. Thus, evolution has occurred due only to random chance – this is genetic drift. While natural selection results from aspects of an organism’s environment exerting “selective pressure” on the individual (e.g., the desert environment favors the spines of the cactus and the long ears of the fox), genetic drift is not a result of environmental pressures.
Genetic drift, like natural selection, is occurring all of the time in all environments. Drift will influence every allele, even those that are being naturally selected. Thus, it can be difficult to determine which process dominates because it is often nearly impossible to determine the cause of change in allele frequencies at any given time.
Genetic drift exerts a particularly strong effect in small populations, such as those that colonize islands or the few individuals that remain after large-scale disruptions (e.g. earthquakes, fire). The statistical principle known as sampling bias underlies this: the random loss of 20 iguanas from a large population of 1 million iguanas is bound to result in a smaller impact on allele frequencies than the random loss of 20 iguanas from an island population of 100 iguanas. Genetic drift occurs because the alleles in an offspring generation are a random sample of the alleles in the parent generation. Alleles may or may not make it into the next generation due to chance events including the mortality of an individual, events affecting finding a mate, and even the events affecting which gametes end up being fertilized. If one individual in a population of ten individuals happens to die before it leaves any offspring to the next generation, all of its genes—a tenth of the population’s gene pool—will be suddenly lost. In a population of 100, that 1 individual represents only 1 percent of the overall gene pool; therefore, it has much less impact on the population’s genetic structure and is unlikely to remove all copies of even a relatively rare allele.
Imagine a population of ten haploid individuals, half with the dominant allele “A” and half with allele the recessive allele “a”. In a stable population, the next generation will also have ten individuals. Choose that generation randomly by flipping a coin ten times and let heads be “A” and tails be “a”. It is unlikely that the next generation will have exactly half of each allele. There might be six of one and four of the other or some different set of frequencies. Thus, the allele frequencies have changed and evolution has occurred. A coin will no longer work to choose the next generation (because the odds are no longer one-half for each allele). The frequency in each generation will drift up and down on what is known as a random walk until at one point either all “A” or all “a” are chosen and that allele is fixed from that point on. This could take a very long time for a large population.
This simplification is not very biological, but simulations and studies have shown that real populations behave this way. The effect of drift on frequencies is greater the smaller a population is. Its effect is also greater on an allele with a frequency far from one-half. Genetic drift causes random changes in allele frequencies and these changes are larger when populations are small.

Bottleneck effect & Founder Effect
Natural events, such as an earthquake disaster that randomly kills a large portion of the population, can magnify the effects of genetic drift. These situations, where a large portion of the gene pool is suddenly and randomly eliminated, is called the bottleneck effect (Fig 1). At once, the survivors’ genetic structure (i.e., allele frequencies) becomes the entire population’s genetic structure, which may be very different from the pre-disaster population. Almost certainly rare alleles are lost from the population, resulting in a decrease in the variability in the population. The survivors reproduce to create the next generation, which can have dramatically different allele frequencies compared to the population before the bottleneck event occurred. Bottleneck effects are a special case of genetic drift where the effects of drift are particularly strong because a population’s size has been dramatically reduced.
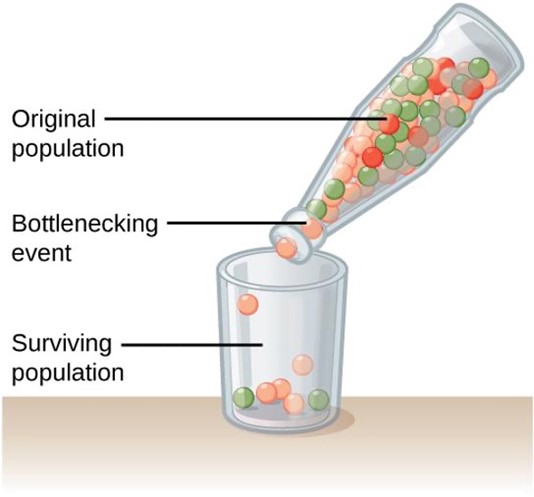
Figure 1: A chance event or catastrophe that dramatically reduces the population size can dramatically alter the allele frequencies in the population.
Another scenario in which populations might experience a strong influence of genetic drift is if some portion of the population leaves to start a new population in a new location. It is likely that the small number of individuals who found the new population have different allele frequencies than the entire population due to random chance (sampling error). Thus, the new population can end up having very different allele frequencies than the original source population due to genetic drift – this is called the founder effect.
Gene Flow / Immigration & Emigration
Gene flow refers to the flow of alleles in and out of a population due to the migration of individuals or gametes (Fig 2). While some populations are fairly stable, others experience more flux. Many plants, for example, send their pollen far and wide, by wind or by a pollinator, to pollinate other populations of the same species some distance away. Even a population that may initially appear to be stable, such as a pride of lions, experiences immigration (coming into) and emigration (departing from) as developing males leave their mothers to seek out a new pride with genetically unrelated females. The flow of individuals in and out of populations changes the population’s allele frequencies. When an individual migrates into a new population, it may sometimes carry alleles that were not present in that population before, introducing new genetic variation into that population (Fig 2). Alternatively, if an individual emigrates from (leaves) a population where it is the only individual with a particular allele, it can result in its original population losing genetic diversity. Migration between populations causes them to become more genetically similar to each other. For example, alleles that may be present in one population but not the other can become present in both populations when a migrating individual introduces that allele to the new population.
In either case, emigration or immigration, if it changes the allelic frequency of a trait results in evolution. For example, if the trait in question is coloration and a population of beetles has only the variation or allele of green color, if a brown beetle immigrates into this population, the allelic frequency has changed. Evolution has occurred (Fig 2).
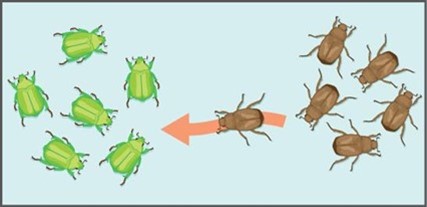
Figure 2: Gene flow occurs when an individual migrates from one population to another. This results in non-adaptive evolution.
There are many examples of populations evolving through gene flow. An appreciation of gene flow can help us understand the global distribution of HIV resistance in humans. The CCR5 mutation confers resistance to some forms of HIV, yet is not most common in areas with a high prevalence of HIV and AIDS. The mutation is relatively new: biochemical and biogeographic evidence suggests an origin in Northern Europe approximately 1,200 years ago. However, the mutation was distributed globally long before HIV and AIDS were relevant to human health. The mutation’s distribution pattern mirrors the Viking migration of the 9th through 11th centuries. Thus, we can hypothesize that Vikings carried the mutation with them as they conquered new territories, and passed the mutation to their descendants. But why was this genetic feature prevalent in Vikings? It could be due to an advantageous mutation. It could have been due to the Founder’s effect.
Mutation
A mutation is a random change in the DNA sequence. There are two types of mutations: point and non-point (sometimes called “silent”). A non-point mutation is a change in the DNA; however, the protein coded by this DNA does not change. There was a change in the DNA but you would not know it from looking at the phenotype. Point mutations are the more powerful types arguably because with point mutations, a change in the DNA results in a change of proteins. In other words, a variation of a trait may be introduced.
Point mutations are the ultimate source of genetic variation in all populations—new alleles, and, therefore, new genetic variations arise through point mutation. Often scientists just use the term mutation to refer to these point mutations. A (point) mutation creates a new allele, and thus by definition results in a small change in the allele frequencies within the population in which it occurs. This means point mutations lead to evolution.
A mutation may produce an allele that is selected against, selected for, or selectively neutral. Harmful mutations (also called deleterious mutations) are removed from the population by selection and will generally only be found in very low frequencies equal to the mutation rate. Some are beneficial and will spread through the population. These are the types of mutations that often we associate with natural selection. If individuals who have the beneficial mutation in a given environment are favored, that is the new phenotype offers adaptation to the selective pressure, they will live longer and reproduce more to pass on this genotype.
Whether a mutation is beneficial or harmful is determined by the environmental context in which it exists. If it helps an organism survive and reproduce in its environment, it is beneficial and therefore an adaptation. Some mutations do not have any impact on fitness (called neutral mutations). The frequency of neutral mutations is unaffected by selection, and only subject to the other mechanisms of evolution.

Questions
Glossary
References
Clark, M.A., Douglas, M., and Choi, J. 2018. Biology 2e. OpenStax.
Fowler, S., Roush, R., & Wise, J. (2022). NSCC Academic Biology 1050. Nova Scotia Community College.
